Свойства нервных центров. Принципы координационной деятельности ЦНС
Координация деятельности нервной системы (взаимодействие ее структур, отдельных рефлексов, программ активности) обеспечивается прохождением информации по строго определенным путям – генетически детерминированным (врожденным) нейронным сетям — специфическим соединениям нейронов.
Классификация нейронных сетей
Генетически детерминированные нейронные сети подразделяют на иерархическую, локальную, дивергентную и конвергентную сеть, реверберирующие нервные цепочки.
Иерархический тип межнейронных сетей имеет несколько уровней переключательных ядер, обеспечивающих очень точную передачу информации. Данные сети наиболее распространены в двигательных и сенсорных путях, а также характерны для сетей, выполняющих какую-то специфическую функцию. В двигательных системах иерархия нисходящая: моторная область коры больших полушарий посылает информацию через переключательные ядра ствола специфическим двигательным клеткам спинного мозга, а те далее «спускают команду» мышечным клеткам. У сенсорных систем иерархическая организация — восходящая.
Локальные сети широко распространены во всех мозговых сетях и удерживают поток информации в пределах одного иерархического уровня, часто действуя как фильтры. Образованные возбуждающими и тормозными интернейронами, они способны преобразовывать импульсы. В сочетании с дивергентными и конвергентными сетями локальные сети позволяют расширять, сужать или снова фокусировать поток информации на определенном иерархическом уровне.
Конвергентные сети обнаружены на всех нервных уровнях и представляют собой взаимодействие возбуждающих и тормозных вставочных интернейронов разных уровней. Мощные конвергентные пути сходятся на нейронах ретикулярной формации, на командных нейронах коры больших полушарий, на мотонейронах спинного мозга. Конвергентные пути, подходящие к одному нейрону, делают этот нейрон интегратором соответствующих сигналов. Данный нейрон-интегратор определяет общий путь для приходящих к нему нервных импульсов.
Дивергентные сети составляют лишь небольшую часть всех нервных сетей (представлены в некоторых ядрах среднего мозга и ствола мозга) и образованы небольшими скоплениями нервных клеток. В данных сетях один нейрон имеет длинный аксон со множеством коллатералей, образующих контакты с огромным числом клеток в различных структурах мозга, и медиатор с «условным» действием (в зависимости от условий эффект от них может меняться). Действие дивергентных путей этих сетей часто не ограничено какой-либо специфической сенсорной, двигательной или другой функциональной системой. Однако, такое неспецифическое влияние на различные иерархические уровни и специфические функции, оказывает важную роль в интегративной деятельности ЦНС, обеспечивая согласованную их работу.
Нервный центр
Объединение сетей между собой образует нервный центр — совокупность нейронов, расположенных в различных отделах ЦНС и обеспечивающих регуляцию определенной специфической функции. Также в регуляции функции могут участвовать несколько нервных центров, дополняющих и дублирующих друг друга. По типу регулируемых функций нервные центры бывают соматические, вегетативные и психические. По своим функциям нервные центры бывают моторные или сенсорные. По локализации различают спинальные, стволовые, мозжечковые, подкорковые и корковые нервные центры.
Свойства нервных центров
Независимо от выполняемой функции, нервные центры обладают рядом общих свойств, обусловленных располагающимися в них телами нейронов и синапсов:
1.Одностороннее проведение возбуждения
В нервном центре возбуждение распространяется в направлении от афферентных нейронов к эфферентным. Это обусловлено односторонним проведением возбуждения в синапсах (выделением медиатора из пресинаптической мембраны).
2. Задержка проведения возбуждения
Через нервные центры возбуждение проводится медленнее, чем по нервам. Происходит центральная или синаптическая задержка из-за наличия в нервных центрах большого числа синапсов. Центральная задержка объясняется затратой времени на выделение из пресинаптической мембраны медиатора, его взаимодействие с рецепторами на постсинаптической мембране, на возникновение ВПСП и формирование потенциала действия на аксонном холмике. Практически все время осуществления рефлекса тратится на проведение возбуждения по соответствующему центру.
Выделяют время рефлекса и центральное время этого рефлекса.
Время рефлекса – это время от начала раздражения рецептора до появления ответной реакции (латентный период).
Центральное время рефлекса – это время, в течение которого происходит внутрицентральное проведение возбуждения. Сила рефлекса зависит от числа активированных эффекторных нейронов (например, мотонейронов). С усилением раздражения рецепторов сила рефлекторного ответа увеличивается до определенного предела.
3. Суммация возбуждений
Она проявляется в усилении рефлекторного ответа при увеличении до определенного предела частоты раздражений (временная или последовательная суммация), либо при увеличении количества возбужденных афферентных нервных волокон или рецепторов (пространственная суммация) (рис. 7.). Основой обоих видов суммации является суммация ВПСП на мембране нейрона. Выделяемые на одиночный импульс кванты медиатора вызывают деполяризацию мембраны, т.е. возникновение ВПСП недостаточного для достижения КУД и развития ПД на аксоном холмике. При временной суммации частые подпороговые стимулы вызывают возникновение ПД в результате суммации ВПСП, возникающих на каждый стимул.
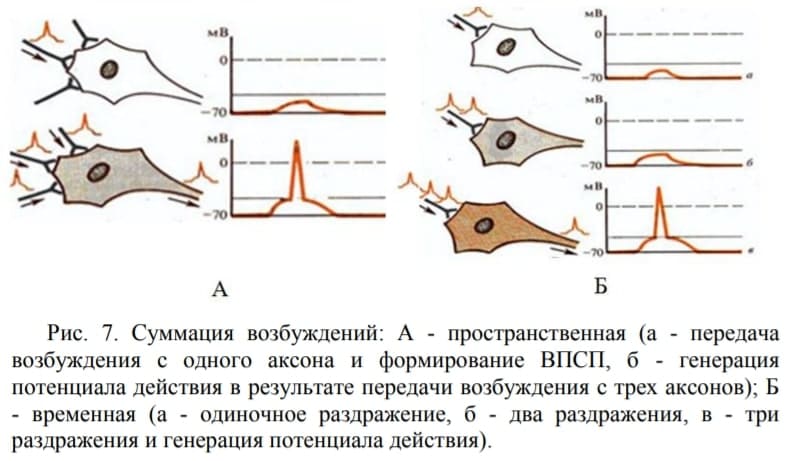
При пространственной суммации рефлекторная реакция наблюдается в результате сложения ВПСП, возникающих от импульсов, одновременно приходящих к нейрону с различных участков одного рецептивного поля. В естественных условиях оба эти вида суммации ВПСП сосуществуют.
4. Облегчение и окклюзия
Пространственная суммация лежит в основе облегчения и окклюзии. Нейроны могут входить в состав разных нервных центров. В каждом же нервном центре выделяют центральную зону и периферическую «кайму». Аксон чувствительного нейрона, приходящий в нервный центр, с помощью терминалей образует множество синаптических контактов с клетками центральной (пороговой) зоны и небольшое количество синапсов в кайме. Таким образом, на нейроны «каймы» приходится незначительная часть импульсов, формирующая подпороговые потенциалы.
Облегчение проявляется в увеличении суммы эффектов при совместном раздражении рецептивных полей двух рефлексов по сравнению с арифметической суммой эффектов при их раздельном раздражении: 8 > 3+3 (рис. 8. А). Наблюдается это, если у нервных центров этих рефлексов имеются общие нейроны, входящие в «каймы»: в результате они получают больше одновременно приходящих импульсов и возбуждаются в результате суммации их подпороговых потенциалов до КУД.
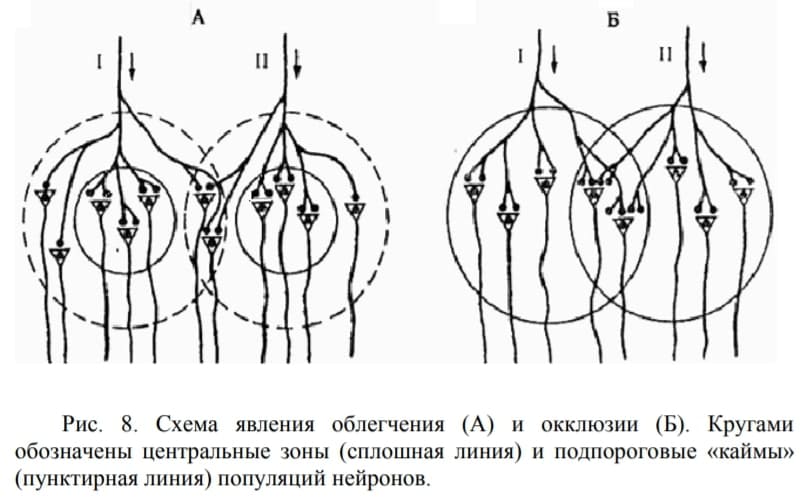
При окклюзии (закупорке) центральные зоны различных нервных центров имеют часть общих нейронов. При совместном раздражении обоих нервных центров суммарный физиологический эффект оказывается меньше, чем арифметическая сумма этих эффектов при изолированном возбуждении каждого из этих центров: 8 < 5+5 (рис. 8. Б). Окклюзия может также наступать при сильном процессе возбуждения, когда общими являются мотонейроны каймы.
5. Посттетаническая потенциация
Посттетаническая потенциация — усиление возбудимости нервного центра после его длительной ритмичной импульсации (с частотой более 300- 600 имп/с).
Посттетаническая потенциация относится к процессам облегчения в ЦНС и имеет отношение к обучению и памяти. Данный эффект объясняется накоплением в пресинаптическом окончании ионов Са2+, а поэтому и большим выделением медиатора. Медиатор выделяется в синаптическую щель в больших количествах, что выражается в увеличении ответной реакции.
6. Трансформация ритма
В нервном центре может меняться частота приходящих импульсов, т. е. частота импульсов на выходе из нервного центра может отличаться от частоты на входе в него. Основным механизмом изменения ритма являются кольцевые нервные сети с положительной и отрицательной обратными связями. В изменении ритма задействованы и мембранные механизмы.
При положительной трансформации ритма — трансформации на увеличение: частота ответной импульсации нервного центра больше, чем частота поступающая от раздражителя. Объясняется это высокой (выше КУД) следовой деполяризацией или длительным ВПСП на постсинаптической мембране нейронов.
При отрицательной трансформации ритма — частота ответной импульсации нервного центра меньше, чем частота поступающая от раздражителя. Объясняется это следовой гиперполяризацией мембраны.
7. Последействие
Свойство проявляется в способности нервного центра сохранять возбуждение еще какое-то время после прекращения импульсации от раздражителя. Как и при трансформации ритма, последействие обусловлено мембранными механизмами — выраженной следовой деполяризацией или возникновением высокоамплитудных, полисинаптических ВПСП в нейронах ЦНС, которые приводят к кратковременному последействию.
Сетевые механизмы — циркуляция возбуждения по замкнутым кольцевым сетям (по вставочным интернейронам) нервных центров, т.е. по нейронным «ловушкам» (рис. 9. В) формируют долговременное последействие, которое может сохраняться достаточно длительное время.
8. Тонус нервных центров
В нервном центре постоянно существует фоновая биоэлектрическая активность, которая обусловлена активирующим влиянием со стороны вышележащих отделов ЦНС и постоянным поступлением импульсов по афферентным волокнам (обратная связь).
9. Утомление нервных центров
Нервные центры легко утомляемы. Это проявляется в частичном или полном прекращении рефлекторного ответа.
Причиной утомления нервных центров является истощение запасов медиатора в нервных окончаниях; уменьшение чувствительности постсинаптической мембраны к его действию, а также уменьшение энергетических запасов нейрона.
10. Высокая чувствительность нервных центров к недостатку кислорода и действию фармакологических веществ
Из-за высокой интенсивности аэробных процессов в нейронах гипоксия приводит к глубокому нарушению деятельности ЦНС. Чувствительность нервного центра к действию фармакологических веществ определяется химическим сродством фармакологических агентов к рецепторам нейронов и позволяет проводить избирательную регуляцию свойств нервного центра.
11. Пластичность нервных центров
При изменении условий нервные центры способны значительно расширять или менять свои функциональные возможности. Пластичность увеличивается в направлении от нервных центров спинного мозга к центрам коры больших полушарий. Способность обучаться, восстанавливаться и переучиваться максимальна в растущем организме и снижается с возрастом.
Конвергенция, дивергенция и реверберация
Проведение возбуждения в ЦНС реализуется за счет конвергенции, дивергенции и реверберации (рис. 9. А).
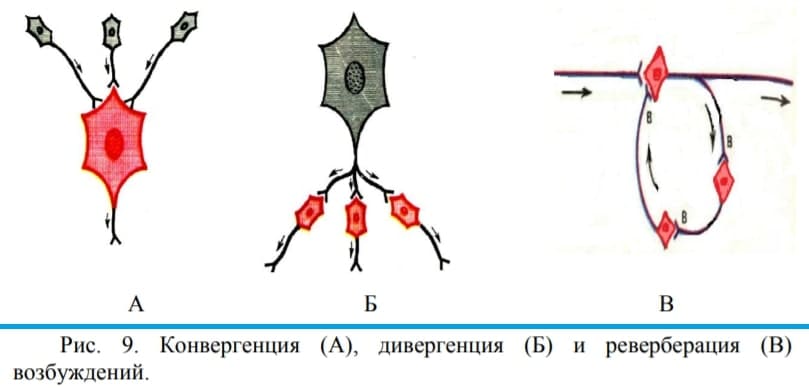
Конвергенция – схождение нескольких нервных сетей на одном нейроне или нервном центре. Благодаря конвергенции происходит «сжатие» объема информации (концентрация), поступающей от рецепторов к ЦНС и формируется интегративный ответ.
Дивергенция — это свойство противоположное конвергенции — «расхождения» путей возбуждения на другие нервные сети (рис. 9. Б). При дивергенции возбуждение от одного нейрона в нервном центре может переключаться на другие нейроны центра. Дивергенция обеспечивает иррадиацию – процесс распространения возбуждения от одного нейрона одновременно на многие другие.
Реверберация — циркуляция импульсов в ЦНС по замкнутым цепочкам нейронов «нейронным ловушкам» (Рис. 9. В.). Циркуляция может продолжаться до тех пор, пока в ней не наступит утомление или какой-либо внешний тормоз не выключит одно из звеньев цепи. Реверберация лежит в основе последействия и кратковременной памяти.
Принципы координации деятельности ЦНС
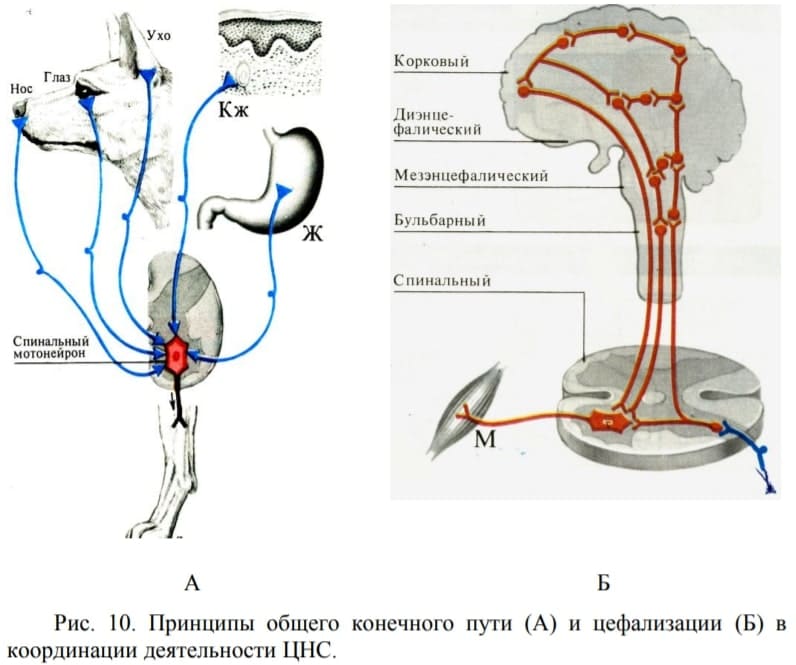
- Принцип общего конечного пути: импульсы, приходящие в ЦНС от разных рецепторов могут сходиться на общих эфферентных нейронах и вызывать одни и те же рефлексы (Рис. 10. А). Один и тот же мотонейрон может включаться в различные рефлекторные дуги. Эти эфферентные нейроны образуют общий конечный путь самых разнообразных рефлексов и могут быть связаны с самыми различными рецепторными аппаратами через вставочные (промежуточные) нейроны.
- Принцип доминанты: рефлексы, реализация которых наиболее важна для организма в данный момент времени, реализуются первыми, а осуществление других (менее значительных) — откладывается (тормозится). Принцип доминанты, который был открыт А.А. Ухтомским, один из основных принципов работы ЦНС, характеризующий наличие в ней «господствующих» центров с высоким уровнем тонической активности, удовлетворяющих жизненно важную потребность.
- Принцип обратной связи. Коррекция большинства рефлексов осуществляется при участии обратной афферентации, необходимой для обратной связи с центром для информирования о достижении ожидаемого результата и координации деятельности.
- Принцип субординации (соподчинения). Принцип иерархичности. В ЦНС регуляция и координация функций характеризуется подчинением низших (филогенетически ранних) отделов высших (филогенетически более поздним). Цефализация заключается в сосредоточении регуляторных и координационных функций в структурах головного мозга (Рис. 10. Б), их высшее проявление – кортикализация — вовлечение в разнообразные формы двигательной активности коры больших полушарий. При прекращении супраспинальных влияний наблюдается обратимое выключение спинальных рефлексов — спинальный шок. После прекращения спинального шока спинальные рефлексы восстанавливаются, однако проводниковая функция спинного мозга – нет.
- Принцип реципрокности (сопряженного взаимодействия). Осуществление противоположных рефлексов (сгибание и разгибание, жевание и глотание, и т.д.) возможно из-за согласованной работы их нервных центров: нейроны одного центра, возбуждаясь, тормозят через вставочные тормозные клетки нейроны другого и наоборот.
- Принцип компенсации функций. Этот принцип основан на способности нервных структур к пластичности – при повреждении отдельных центров, их функция компенсируются за счет других центров и коры больших полушарий мозга.