Биохимия
Классификация белков
По функции
По строению
1. По форме молекулы
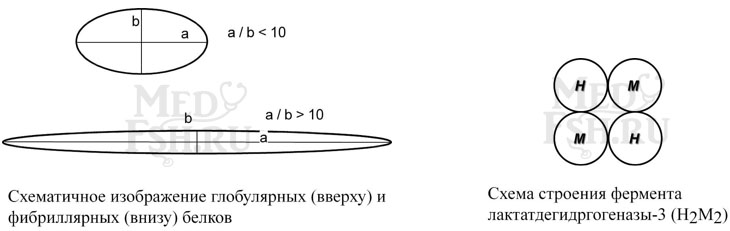
Глобулярные – соотношение продольной и поперечной осей составляет <10 и в большинстве случаев не более 3-4. Они характеризуются компактной укладкой полипептидных цепей. Например, инсулин, альбумин, глобулины плазмы крови.
Фибриллярные – соотношение осей >10. Они состоят из пучков полипептидных цепей, спиралью навитых друг на друга и связанные между собой поперечными ковалентными и водородными связями. Выполняют защитную и структурную функции. Например, кератин, миозин, коллаген.
2. По количеству белковых цепей в одной молекуле
- мономерные – имеют одну субъединицу (протомер), например, альбумин, миоглобин.
- полимерные – имеют несколько субъединиц. Например, гемоглобин (4 субъединицы), лактатдегидрогеназа (4 субъединицы), креатинкиназа (2 субъединицы)
3. По химическому составу
Простые – содержат в составе только аминокислоты (альбумины, глобулины, гистоны, протамины). Подробно эти белки характеризуются ниже.
Сложные – кроме аминокислот имеются небелковые компоненты (нуклеопротеины, фосфопротеины, металлопротеины, липопротеины, хромопротеины, гликопротеины). Подробно эти белки характеризуются ниже.
Классификация простых белков
Структура простых белков представлена только полипептидной цепью (альбумин, инсулин). Однако необходимо понимать, что многие простые белки (например, альбумин) не существуют в „чистом» виде, они всегда связаны с какими-либо небелковыми веществами. Их относят к простым белкам, т.к. связи с небелковой группой слабые.
Альбумины
Группа глобулярных белков плазмы крови с молекулярной массой 69 кДа, имеют кислые свойства и отрицательный заряд при физиологических рН, т.к. содержат много глутаминовой кислоты. Легко адсорбируют полярные и неполярные молекулы, являются в крови переносчиком многих веществ, в первую очередь билирубина и жирных кислот.
В природе альбумины входят в состав не только плазмы крови (сывороточные альбумины), но и яичного белка (овальбумин), молока (лактальбумин), являются запасными белками семян высших растений.
Глобулины
Группа разнообразных белков плазмы крови с молекулярной массой до 100 кДа, несут отрицательный заряд, слабокислые или нейтральные. Они мало гидратированы, по сравнению с альбуминами меньше устойчивы в растворе и легче осаждаются, что используется в клинической диагностике в «осадочных» пробах (тимоловая проба, проба Вельтмана). Часто содержат углеводные компоненты.
При обычном электрофорезе разделяются, как минимум, на 4 фракции – α1, α 2, β и γ.
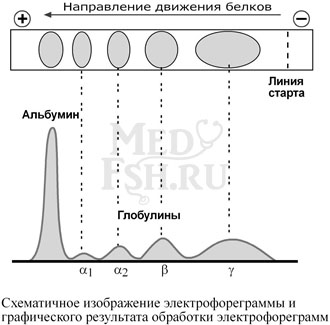
Так как глобулины включают в себя разнообразные белки, то их функции многочисленны. Часть α-глобулинов обладает антипротеазной активностью, что защищает белки крови от преждевременного разрушения, например, α 1-антитрипсин, α 1-антихимотрипсин, α 2-макроглобулин. Некоторые глобулины способны к связыванию определенных веществ: трансферрин (переносчик ионов железа), церулоплазмин (переносчик ионов меди), гаптоглобин (переносчик гемоглобина), гемопексин (переносчик гема). γ -глобулины являются антителами и обеспечивают иммунную защиту организма.
Гистоны
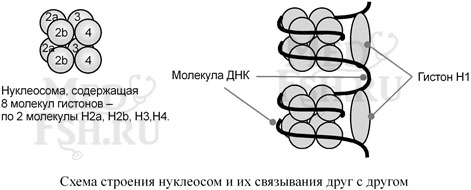
Гистоны – внутриядерные белки массой около 24 кДа. Обладают выраженными основными свойствами, поэтому при физиологических значениях рН заряжены положительно и связываются с дезоксирибонуклеиновой кислотой (ДНК), образуя дезоксирибонуклеопротеины. Существуют 5 типов гистонов – очень богатый лизином (29%) гистон Н1, другие гистоны Н2а, H2b, НЗ, Н4 богаты лизином и аргинином (в сумме до 25%).
Радикалы аминокислот в составе гистонов могут быть метилированы, ацетилированы или фосфорилированы. Это изменяет суммарный заряд и другие свойства белков.
Можно выделить две функции гистонов:
- Регулируют активность генома, а именно препятствуют транскрипции.
- Структурная – стабилизируют пространственную структуру ДНК.
Гистоны образуют нуклеосомы – октаэдрические структуры, составленные из гистонов Н2а, H2b, НЗ, Н4. Гистон H1 связан с молекулой ДНК, не позволяя ей «соскользнуть» с гистонового октамера. ДНК обвивает нуклеосому 2,5 раза после чего обвивает следующую нуклеосому. Благодаря такой структуре достигается уменьшение размеров ДНК в 7 раз. Далее нить ДНК с нуклеосомами складывается в суперспираль и «суперсуперспираль». Таким образом, гистоны участвуют в плотной упаковке ДНК при формировании хромосом.
Благодаря гистонам в конечном итоге размеры ДНК уменьшаются в тысячи раз: длина ДНК достигает 6-9 см (10–1), а размеры хромосом – всего несколько микрометров (10–6).
Протамины
Это белки массой от 4 кДа до 12 кДа, имеются в ядрах сперматозоидов многих организмов, в сперме рыб они составляют основную массу белка. Протамины являются заменителями гистонов и служат для организации хроматина в спермиях. По сравнению с гистонами протамины отличаются резко увеличенным содержанием аргинина (до 80%). Также, в отличие от гистонов, протамины обладают только структурной функцией, регулирующей функции у них нет, хроматин в сперматозоидах неактивен.
Коллаген
Фибриллярный белок с уникальной структурой. Составляет основу межклеточного вещества соединительной ткани сухожилий, кости, хряща, кожи, но имеется, конечно, и в других тканях.
Полипептидная цепь коллагена включает 1000 аминокислот и носит название α-цепь. Насчитывается около 30 вариантов α-цепи коллагена, но все они обладают одним общим признаком – в большей или меньшей степени включают повторяющийся триплет [Гли-Х-Y], где X и Y – любые, кроме глицина, аминокислоты. В положении X чаще находится пролин или, гораздо реже, 3-оксипролин, в положении Y встречается пролин и 4-оксипролин. Также в положении Y часто находится аланин, лизин и 5-оксилизин. На другие аминокислоты приходится около трети от всего количества аминокислот. Гидроксилирование пролина и лизина осуществляют соответствующие гидроксилазы, железосодержащие ферменты, для их полноценной работы необходим витамин С (аскорбиновая кислота).
Структура пролина и оксипролина не позволяет образовать α-спиральную структуру, из-за этого образуется левозакрученная спираль, где на один виток приходится 3 аминокислотных остатка. Коллаген обычно содержит моносахаридные (галактоза) и дисахаридные (глюкоза-галактоза) молекулы, соединенные с ОН-группами некоторых остатков оксилизина.
Молекула тропоколлагена построена из 3 α-цепей, сплетенных между собой в плотный жгут – тропоколлаген (длина 300 нм, диаметр 1,6 нм). Полипептидные цепи прочно связаны между собой через ε-аминогруппы (ε-эпсилон) остатков лизина. Тропоколлаген формирует крупные коллагеновые фибриллы диаметром 10-300 нм. Поперечная исчерченность фибриллы обусловлена смещением молекул тропоколлагена друг относительно друга на 1/4 их длины.

Время полужизни коллагена исчисляется неделями и месяцами. Ключевую роль в его обмене играет коллагеназа, расщепляющая тропоколлаген на 1/4 расстояния с С-конца между глицином и лейцином. В результате распада коллагена появляется свободный гидроксипролин. При поражении соединительной ткани (болезнь Пейджета, гиперпаратиреоидизм) экскреция гидроксипролина возрастает и имеет диагностическое значение.
По мере старения организма в тропоколлагене образуется все большее число поперечных связей, что делает фибриллы коллагена в соединительной ткани более жесткими и хрупкими. Это ведет к повышенной ломкости кости и снижению прозрачности роговицы глаза в старческом возрасте.
Эластин
По строению в общих чертах эластин схож с коллагеном. Находится в связках, эластичном слое сосудов. Структурной единицей является тропоэластин с молекулярной массой 72 кДа и длиной 800 аминокислотных остатков. В нем гораздо больше лизина, валина, аланина и меньше гидроксипролина. Отсутствие пролина обусловливает наличие спиральных эластичных участков.
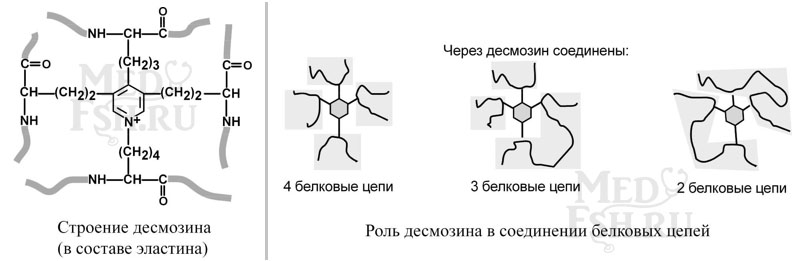
Характерной особенностью эластина является формирование из четырех молекул лизина своеобразной структуры – десмозина, который своими 4-мя группами объединяет белковые цепи в системы, способные растягиваться во всех направлениях.
α-аминогруппы и α-карбоксильные группы десмозина включаются в пептидные связи одной или нескольких белковых цепей.
Строение и классификация сложных белков
У сложных белков, кроме белковой цепи, имеется дополнительная небелковая группа. Она называется лиганд, (лат. ligo – связываю) то есть молекула, связанная с белком. В случае если лиганд несет структурную и/или функциональную нагрузку, он называется простетической группой.
В роли лиганда могут выступать любые молекулы:
- молекулы, выполняющие в белке структурную функцию – липиды, углеводы, нуклеиновые кислоты, минеральные элементы, какие-либо другие органические соединения: гем в гемоглобине, углеводы в гликопротеинах, ДНК и РНК в нуклеопротеинах, медь в церулоплазмине,
- переносимые белками молекулы: железо в трансферрине, гемоглобин в гаптоглобине, гем в гемопексине,
- субстраты для ферментов – любые молекулы и даже другие белки.
Узнавание лиганда обеспечивается:
- комплементарностью структуры центра связывания белка структуре лиганда, иначе говоря, пространственным и химическим соответствием белка и лиганда. Они подходят друг к другу как ключ к замку, например, соответствие фермента и субстрата,
- иногда узнавание может зависеть от реакционной способности атома, к которому присоединяется лиганд. Например, связывание кислорода железом гемоглобина, или жирной кислоты с альбумином.
Функции лиганда в составе сложного белка разнообразны:
- изменяет свойства белков (заряд, растворимость, термолабильность), например, фосфорная кислота в фосфопротеинах или остатки моносахаридов в гликопротеинах,
- защищает белок от протеолиза вне и внутри клетки, например, углеводная часть в гликопротеинах,
- обеспечивает возможность транспорта нерастворимых в воде соединений, например, перенос жиров липопротеинами,
- придает биологическую активность и определяет функцию белка, например, нуклеиновая кислота в нуклеопротеинах, гем в гемоглобине, углевод в рецепторных белках,
- влияет на проникновение через мембраны, внутриклеточную миграцию, сортировку и секрецию белков, это выполняет, как правило, углеводный остаток.
Фосфопротеины
Фосфопротеины – это белки, в которых присутствует фосфатная группа, она связывается с пептидной цепью через остатки тирозина, серина, треонина (т.е. тех аминокислот, которые содержат ОН-группу).
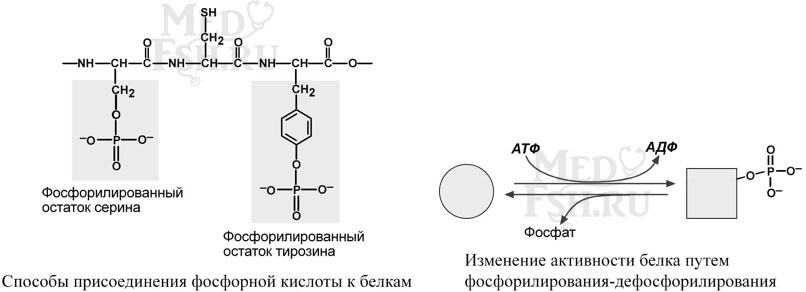
Фосфорная кислота может выполнять:
- Структурную роль, придавая заряд, растворимость и изменяя свойства белка, например, в казеине молока, яичном альбумине.
- Функциональную роль. В клетке присутствует много белков, которые связаны с фосфатом не постоянно, а в зависимости от активности метаболизма. Белок может многократно переходить в фосфорилированную или в дефосфорилированную форму, что играет регулирующую роль в его работе. Например, 1) ферменты гликогенсинтаза и гликогенфосфорилаза (см «Регуляция активности ферментов»), 2) гистоны в фосфорилированном состоянии менее прочно связываются с ДНК и активность генома возрастает.
Нуклеопротеины
Нуклеопротеины – это белки, связанные с нуклеиновыми кислотами. Они составляют существенную часть рибосом, хроматина, вирусов.
В рибосомах рибонуклеиновая кислота (РНК) связывается со специфическими рибосомальными белками.
Вирусы являются практически чистыми рибо- и дезоксирибонуклеопротеинами.
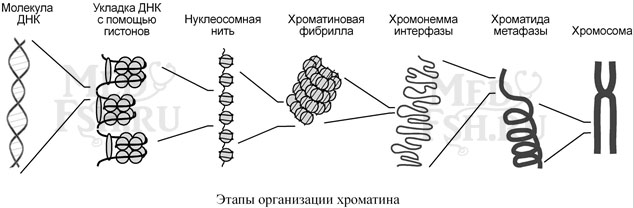
В хроматине нуклеиновая кислота представлена дезоксирибонуклеиновой кислотой, связанной с разнообразными белками, среди которых можно выделить две основные группы – гистоны и негистоновые белки. Начальный этап упаковки ДНК осуществляют гистоны, более высокие уровни обеспечиваются другими белками.
В начале молекула ДНК обвивается вокруг гистонов, образуя нуклеосомы. Сформированная таким образом нуклеосомная нить напоминает бусы, которые складываются в суперспираль (хроматиновая фибрилла) и суперсуперспираль (хромонемма интерфазы).
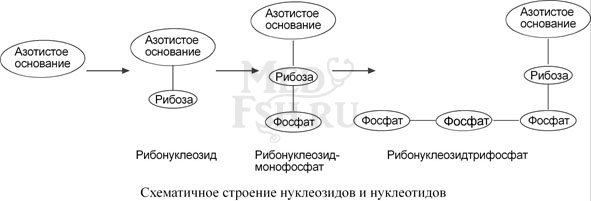
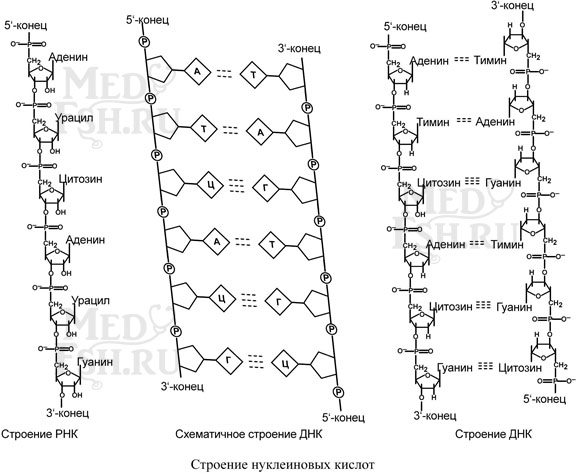
Нуклеиновые кислоты являются полимерными молекулами и состоят из мономеров, называемых нуклеотидами. Нуклеотид содержит фосфорную кислоту (один, два или три остатка), сахар (рибозу или дезоксирибозу), азотистое основание (аденин, гуанин, цитозин, урацил либо тимин).
Самым распространенным в природе является нуклеотид аденозинтрифосфат, используемый как высокоэнергетическое соединение для реакций трансмембранного переноса веществ и реакций синтеза.
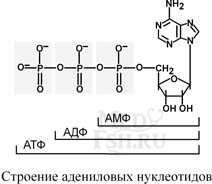
Связываясь через фосфатные остатки, нуклеотиды образуют длинные цепочки – нуклеиновые кислоты.
Выделяют два вида нуклеиновых кислот в зависимости от пентозы, входящей в их состав – рибонуклеиновая кислота (РНК) и дезоксирибонуклеиновая кислота (ДНК). Сахарофосфатный остов в ДНК и РНК заряжен отрицательно благодаря заряду фосфатных групп. В то же время пуриновые и пиримидиновые основания гидрофобны.
Цепи ДНК и РНК обладают направленностью, т.е. имеют 3′-конец и 5′-конец. В ДНК цепи антипараллельны, т.е. направлены в разные стороны. Имеется комплементарность азотистых оснований, соответствие аденин комплементарен тимину (А=Т), гуанин комплементарен цитозину (Г≡Ц).
Липопротеины
К липопротеинам, строго говоря, принадлежат только белки, содержащие ковалентно связанные липиды. Однако традиционно к липопротеинам относят и надмолекулярные образования, выполняющие транспортную функцию и состоящие из белков и молекул всех классов липидов – транспортные липопротеины.
Структуру транспортных липопротеинов можно сравнить с орехом, у которых имеется скорлупа и ядро. «Скорлупа» липопротеина является гидрофильной, ядро – гидрофобное. Ядро формируют неполярные эфиры холестерола и триацилглицеролы. В поверхностном слое («скорлупе») находятся фосфолипиды, свободный холестерол, белки.
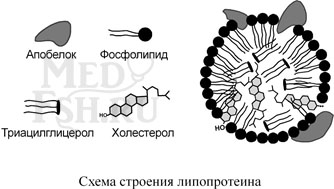
Белки в липопротеинах называются апобелками, их выделяют несколько видов: А, В, С и D. В каждом типе липопротеинов преобладают соответствующие ему апобелки.
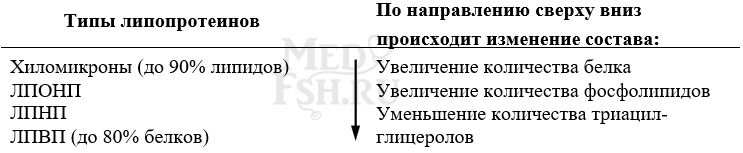
Выделяют четыре основных класса липопротеинов:
- хиломикроны (ХМ),
- липопротеины высокой плотности (ЛПВП, α-липопротеины, α-ЛП),
- липопротеины низкой плотности (ЛПНП, β-липопротеины, β-ЛП),
- липопротеины очень низкой плотности (ЛПОНП, пре-β-липопротеины, пре-β-ЛП).
Липопротеины различаются по составу, т.е. по соотношению триацилглицеролов, холестерола и его эфиров, фосфолипидов, белков.
Хромопротеины
Хромопротеины содержат окрашенные простетические группы. Сюда относят гемопро- теины (содержат гем), ретинальпротеины (содержат витамин А), флавопротеины (содержат витамин В2), кобамидпротеины (содержат витамин В12).
Гемопротеины
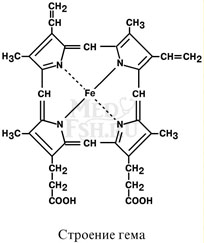
Подразделяются на неферментативные (гемоглобин, миоглобин) и ферменты (цитохромы, каталаза, пероксида- за). Небелковой частью их является гем – структура, включающая в себя порфириновое кольцо (состоящее из 4 пиррольных колец) и иона Fe2+. Железо связывается с порфириновым кольцом двумя координационными и двумя ковалентными связями.
Цитохромы
Цитохромы отличаются аминокислотным составом пептидных цепей и числом цепей и разделяются на типы а, b, с, d. Все они неспособны связывать кислород, кроме цитохрома а3, который содержит ионы меди. Цитохромы находятся в составе дыхательной цепи митохондрий и цепи микросомального окисления.
Флавопротеины
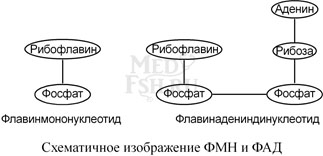
Флавопротеины являются ферментами окислительно-восстановительных реакций, например, сукцинатдегидрогеназа, глицерол-3-фосфатдегидрогеназа. Они в качестве небелковой части содержат производные витамина В2 – флавинмононуклеотид (ФМН) и флавинадениндинуклеотид (ФАД). Флавинадениндинуклеотид представляет комплекс из двух нуклеотидов – аденозинмонофосфата и флавинмононуклеотида.
Металлопротеины
Металлопротеины, кроме белка, содержат ионы одного или нескольких металлов. Металлопротеины часто являются ферментами. Ионы металлов соединены координационными связями с функциональными группами белка и выполняют следующие функции:
- участвуют в ориентации субстрата в активном центре фермента,
- входят в состав активного центра фермента и участвуют в катализе, например, служат акцепторами электронов на определенной стадии ферментативной реакции.
К металлопротеинам, например, относятся белки, содержащие:
- медь – цитохромоксидаза, в комплексе с другими ферментами дыхательной цепи митохондрий участвует в синтезе АТФ,
- железо – ферритин, депонирующий железо в клетке, трансферрин, переносящий железо в крови, сукцинатдегидрогеназа (фермент ЦТК),
- цинк – алкогольдегидрогеназа, обеспечивающая метаболизм этанола и других спиртов, лактатдегидрогеназа, участвующая в метаболизме молочной кислоты, карбоангидраза, образующая угольную кислоту из CO2 и H2O, щелочная фосфатаза, гидролизующая фосфорные эфиры различных соединений.
- селен – тиреопероксидаза, участвующая в синтезе гормонов щитовидной железы, антиоксидантный фермент глутатионпероксидаза,
- кальций – α-амилаза слюны и панкреатического сока, гидролизующая крахмал.
Гликопротеины (гликоконъюгаты)
Гликопротеины – белки, содержащие углеводный компонент, ковалентно присоединенный к полипептидной основе. Содержание углеводов варьирует от 1 до 85% по массе.
Выделяют два подкласса белков, содержащих углеводы: протеогликаны и гликопротеины. Между этими подклассами имеются существенные отличия:
1. Гликопротеины:
- доля углеводов 15-20%,
- не содержат уроновых кислот,
- углеводные цепи содержат не более 15 звеньев,
- углевод имеет нерегулярное строение.
2. Протеогликаны:
- доля углеводов 80-85%,
- имеются уроновые кислоты,
- углеводные цепи крайне велики,
- углевод имеет регулярное строение.
Гликопротеины
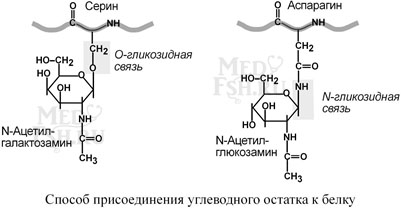
Для собственно гликопротеинов характерно низкое содержание углеводов. Углеводный остаток является олигосахаридом, имеет нерегулярное строение и содержит маннозу, галактозу, глюкозу, и их аминопроизводные, также N-ацетилнейраминовую кислоту.
Олигосахарид присоединен к аминокислотам белковой цепи либо N-гликозидной связью – к амидному азоту аспарагина, либо О-гликозидной связью – к гидроксигруппе остатков серина, треонина, гидроксилизина.
Функцией гликопротеинов являются:
- Структурная – клеточная стенка бактерий, костный матрикс, например, коллаген, эластин.
- Защитная – например, антитела, интерферон, факторы свертывания крови (протромбин, фибриноген).
- Транспортная – перенос веществ в крови и через мембраны, например, трансферрин, транскортин, альбумин, Na+,К+-АТФаза.
- Гормональная – гонадотропный, адренокортикотропный и тиреотропный гормоны.
- Ферментативная – холинэстераза, нуклеаза.
- Рецепторная – присоединение эффектора приводит к изменению конформации белка-рецептора, что вызывает внутриклеточный ответ.
Протеогликаны
Другая группа гликопротеинов – протеогликаны – характеризуется наличием полисахаридов, состоящих из повторяющихся дисахаридных остатков.
Дисахариды включают в себя уроновую кислоту и аминосахар. Многократно дублируясь, дисахариды образуют олиго- и полисахаридные цепи – гликаны. В литературе встречаются другие названия – кислые гетерополисахариды (т.к. имеют много кислотных групп), гликозаминогликаны (содержат аминогруппы). Эти молекулы входят в состав протеогликанов – сложных белков, функцией которых является заполнение межклеточного пространства и удержание здесь воды, также они выступают как смазочный и структурный компонент суставов и других тканевых структур.
Углеводная часть, аналогично с гликопротеинами, связывается с белком через остатки серина и аспарагина.
Основными представителями структурных гликозаминогликанов является гиалуроновая кислота, хондроитинсульфаты, кератансульфаты и дерматансульфаты. Гепарин используется в антикоагулянтной системе крови.
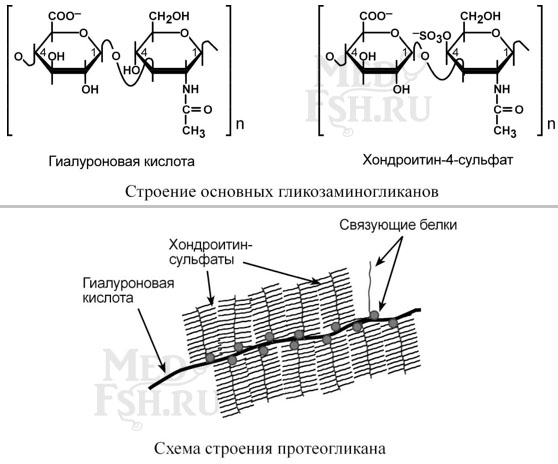
По функции протеогликаны особенно значимы для межклеточного пространства, особенно соединительной ткани, в которое погружены коллагеновые волокна. При помощи электронной микроскопии выяснено, что они имеют древовидную структуру – в центре находится гиалуроновая кислота, которая через связующие белки присоединяет многочисленные «веточки» из хондроитинсульфатов, кератансульфатов, дерматансульфатов. Молекулы гликанов весьма гидрофильны, создают сетчатый желеподобный матрикс и заполняют пространство между клетками, являясь преградой для крупных молекул и микроорганизмов.