Регуляция дыхания
Регуляция дыхания осуществляется ЦНС.
Спокойное дыхание взрослого человека характеризуется последовательной сменой актов вдоха и выдоха с частотой 14-16 дыханий в минуту.
Чтобы произошел вдох, необходимо сокращение дыхательных мышц.
Импульсы к ним поступают от мотонейронов передних рогов спинного мозга.
К диафрагме – от III – IV шейных сегментов, к межреберным мышцам – от грудных сегментов спинного мозга.
Мотонейроны получают импульсы от дыхательного центра, расположенного в продолговатом мозге.
Если перерезать спинной мозг под продолговатым мозгом, то дыхание – прекращается.
Дыхательный центр
Был открыт в 1812 г. Легаллуа и в 1842 г. Флурансом, которые своими опытами доказали его локализацию в продолговатом мозге.
Н.А.Миславский в 1885 г. уточнил местоположение дыхательного центра – в области РФ (ретикулярной формации) продолговатого мозга, т.к. перерезка между грудными и шейными сегментами спинного мозга – сохраняет диафрагмальное дыхание:
- продолговатым и спинным – полностью прекращает дыхание,
- выше продолговатого мозга – дыхание сохраняется.
Н.А.Миславским также было показано, что дыхательный центр состоит из двух отделов:
- инспираторного (отвечает на вдох),
- экспираторного (отвечает за выдох).
Они находятся в реципрокных (то есть в противоположных) отношениях.
В настоящее время установлено:
- в РФ продолговатого мозга находятся инспираторные и экспираторные нейроны,
- эти нейроны рассеяны диффузно, среди других нейронов РФ и относительно друг друга,
- инспираторных нейронов примерно вдвое больше, чем экспираторных.
В продолговатом мозге есть еще два скопления дыхательных нейронов – дорсальные и вентральные дыхательные ядра.
Дорсальное ядро содержит, в основном, инспираторные нейроны, контролирующие сокращение диафрагмы.
В вентральном ядре содержатся как инспираторные, так и экспираторные нейроны, которые включаются при форсированном дыхании.
Все дыхательные нейроны делят на 6 групп:
- ранние инспираторные – активны в начале фазы вдоха (инспирации),
- поздние инспираторные – активны в конце вдоха,
- полные инспираторные – активны в течение всего вдоха,
- постинспираторные – максимальный разряд в начале выдоха,
- экспираторные – активны во вторую фазу выдоха,
- преинспираторные – активны перед вдохом, они включают активную экспирацию (выдох.
Значение дыхательных нейронов:
- Преинспираторные и ранние инспираторные влияют на момент начала вдоха.
- Инспираторные нейроны ДЦ генерируют дыхательный ритм (частоту и глубину дыхания), иннервируют мотонейроны.
- Постинспираторные нейроны контролируют процесс пассивного выдоха.
- Экспираторные нейроны отвечают за активный выдох, так как иннервируют мотонейроны внутренних межреберных мышц и мышц передней брюшной стенки.
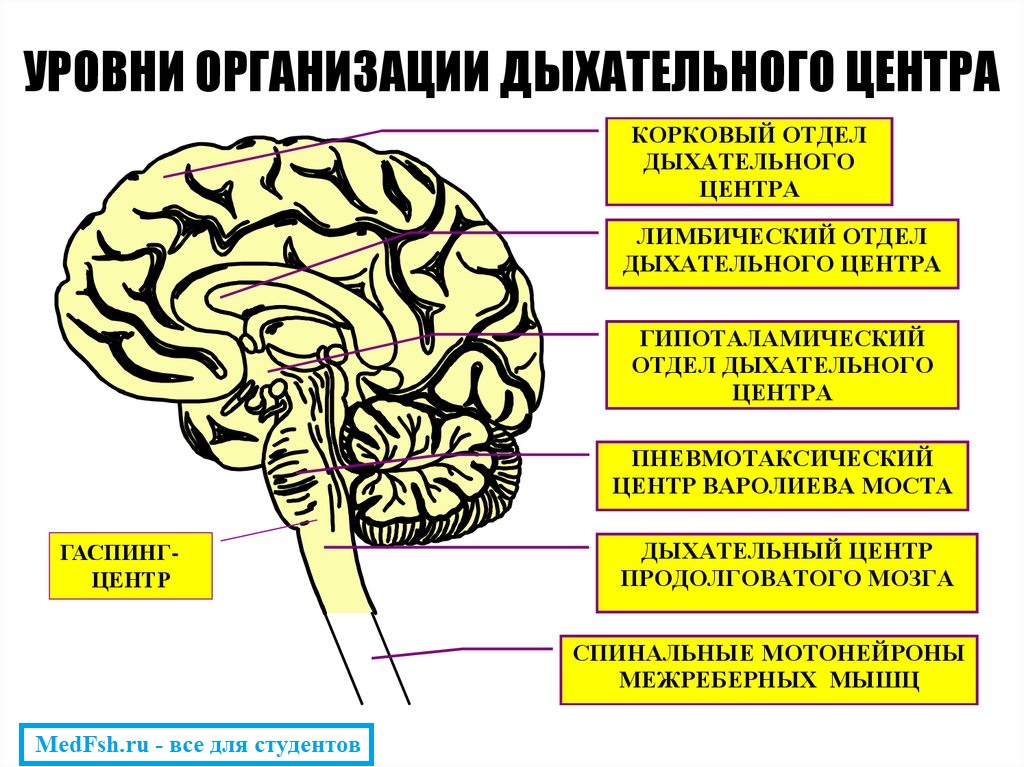
Дыхательный центр – это совокупность нервных клеток, расположенных в различных отделах ЦНС и принимающих участие в регуляции дыхания.
Опыт Фредерика (1901 г.) с перекрестным кровообращением у собак
У двух собак, находящихся под наркозом, перекрестно соединяли сонные артерии и яремные вены, латерально же расположенные сосуды пережимали. При этом голова 1-й собаки снабжались кровью из туловища 2-й и наоборот.
У первой собаки кратковременно пережимали трахею, и у нее в крови уменьшалось содержание кислорода (гипоксемия) и увеличивалось содержание углекислого газа (гиперкапния). Эта кровь поступала в голову 2-й собаки, и у нее наступала одышка (диспноэ).
В результате у нее в крови увеличивалось содержание кислорода (гипероксимия) и уменьшалось содержание углекислого газа (гипокапния), и эта кровь поступала в голову 1-й собаки, и у нее наступало апноэ – остановка дыхания. (Нормальное дыхание — эйпноэ).
На состояние ДЦ влияет газовый состав крови:
При увеличении в крови напряжения углекислого газа и уменьшении кислорода, ДЦ – возбуждается и, наоборот, уменьшается, если в крови уменьшается напряжение углекислого газа и увеличивается напряжение кислорода.
Опыт Холдена
При дыхании в герметичной камере в воздухе увеличивалось содержание CO2 и уменьшалось O2 – наступала одышка. Когда CO2 поглощается натронной известью, одышка наступала намного позже, хотя содержание O2 в воздухе значительно снижалось.
Одышка наступает:
- При снижении O2 в атмосферном воздухе с 20,94% до 12%, т.е. на 9%.
- При повышении содержания CO2 в альвеолярном воздухе на 0,17% вентиляция удваивается.
Главный активатор ДЦ – CO2
Действие карбогена
Карбоген – газовая смесь, состоящая из 96% — O2, 4% — CO2.
В сравнении с воздухом в карбогене в 4,8 раза больше O2 и в 130 раз – CO2.
Карбоген применяют при расстройствах дыхания.
Эффект карбогена связан с эффектами содержащегося в нем CO2:
- стимуляция ДЦ,
- расширение бронхов и кровеносных сосудов,
- сдвиг кривой диссоциации HbO2 вправо –> увеличение диффузии O2 из крови в ткани.
Карбоген применяется в медицинских целях для лечения горной болезни, отравления угарным газом, глаукомы, стресса, при восстановлении слуха после воздействия шума и в ряде случаев для улучшения кровоснабжения опухолей при химио- и лекарственной терапии.
В 1911 г. Винтерштейн доказал, что возбудителем ДЦ также являются ионы Н.
Ацидоз – усиливает легочную вентиляцию.
Он показал, что возбуждают ДЦ нелетучие кислоты – молочная, никотиновая и другие.
Самым сильным стимулятором дыхания являются:
- pCO2 (гиперкапния),
- pH (ацидоз),
- pO2 (гипоксемия).
Механизм действия гуморальных факторов (CO2, O2, H) на ДЦ:
- Через хеморецепторы (периферические) сосудистых рефлексогенных зон.
- Через хеморецепторы, находящиеся в продолговатом мозге (медуллярные).
Периферические хеморецепторы:
- расположены в каротидных и в аортальных тельцах,
- реагируют на (в артериальной крови):
- увеличение pCO2,
- уменьшение pO2,
- увеличение H (от есть уменьшение pH).
Хеморецепторы возбуждаются постоянно CO2 и O2 , растворенными в крови, а также H, т.к. порог для pCO2 равен 20-30 мм.рт.ст. В норме pCO2 = 40 мм.рт.ст.
Порог для pO2 равен 130-140 мм.рт.ст. В норме pO2 = 100 мм.рт.ст. Одышка же наступает при pO2 ниже 50-60 мм.рт.ст.
Таким образом, хеморецепторы постоянно посылают импульсы в ДЦ, возбуждая инспираторные нейроны, причем большую роль играют хеморецепторы каротидного синуса.
Медуллярные хеморецепторы:
- Находятся на вентролатеральной поверхности продолговатого мозга.
- Реагируют только на H и изменение напряжения CO2.
- Эти рецепторы возбуждаются позднее, поскольку требуется время для проникновения CO2 через гематоэнцефалический барьер.
- Импульсы, поступающие с медуллярных хеморецепторов в ДЦ, увеличивают прирост вентиляции на 60-80%.
Механорецепторы (рецепторы растяжения)
Находятся в:
- легких,
- дыхательных путях,
- дыхательных мышцах (проприорецепторы)
Это барорецепторы рефлексогенных зон.
Механорецепторы легких (РРЛ)
Рецепторы растяжения легких (РРЛ) – являются наиболее значимым среди всех механорецепторов.
В 1868 г. Геринг и Брейер доказали наличие в легких рецепторов, которые возбуждаются при их растяжении, то есть при вдохе. Они являются чувствительными окончаниями блуждающих нервов, которые направляют свои импульсы в ДЦ.
Геринг и Брейтер раздували легкие и наблюдали прекражение вдоха (инспираторно-тормозной рефлекс).
Таким образом, этот рефлекс способствует смене вдоха на выдох. Он называется рефлексом Геринга-Брейера и является рефлексом саморегуляции дыхания.
При перерезке блуждающих нервов, дыхание становится редким и глубоким, альвеолы расширяются до максимального предела, т.к. вдох не тормозится. В этом случае, смене вдоха на выдох будет способствовать пневмотаксический центр (ПТЦ).
В настоящее время известно, что в легких существует 3 разновидности механорецепторов:
- РРЛ медленноадаптирующиеся,
- РРЛ быстроадаптирующиеся или ирритантные,
- Юкстаальвеолярные рецепторы капилляров.
РРЛ медленноадаптирующиеся:
- Они расположены в ГМК (гладкомышечных клетках) дыхательных путей.
- Возбуждаются при вдохе.
- С них осуществляется рефлекс Геринга-Брейера.
Ирритантные рецепторы (быстроадаптирующиеся):
- Расположены в слизистой дыхательных путей.
- Реагируют на механические и химические стимулы.
- Быстро адаптирующиеся.
- Длительное раздражение этих рецепторов приводит к хроническому бронхиту.
Физиологическое значение при вдыхании токсических веществ: Сужение бронхов -> вентиляция альвеол -> поступление этих веществ в альвеолы и кровь.
Юкстаальвеолярные рецепторы («юкстакапиллярные»):
- расположены в паренхиме легких, в альвеолярных перегородках, прилегающих к капиллярам,
- стимулируются, главным образом, растяжением легочных сосудов,
- быстро реагируют на введение химических веществ в легочные сосуды,
- стимуляция может вызвать апноэ, затем учащение дыхания, уменьшение давления, брадикардию и бронхоспазм.
Возбуждение механорецепторов верхних ДП вызывает возникновение дыхательных рефлексов.
Защитные дыхательные рефлексы:
- Чихание – с рецепторов слизистой носа.
- Кашель – с ирритантных рецепторов слизистой гортани, трахеи, бронхов.
- Рефлекс ныряльщиков – остановка дыхания при действии воды на носовые ходы.
- Остановка дыхания во время акта глотания.
- Рефлексогенная задержка дыхания – сужение голосовой щели, бронхоконстрикция при вдыхании дыма, газов, едких веществ.
Механорецепторы межреберных мышц и диафрагмы (проприорецепторы) – они осуществляют обратную связь дыхательных мышц с ДЦ.
Барорецепторы рефлексогенных зон – возбуждаются они при увеличении АД и дыхание при этом угнетается. При уменьшении АД – дыхание усиливается.
Ритмическая смена дыхательных фаз
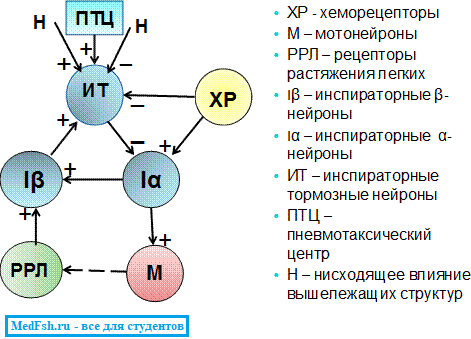
- Генератор центральной инспираторной активности (возбуждения) – сокр. ЦИА (В) – представлен α-инспираторными нейронами дорсального ядра. Они возбуждаются от хеморецепторов (центральных и сосудистых рефлексогенных зон). Чем больше раздражение хеморецепторов (ХР), тем больше скорость ЦИА.
- Механизм выключения инспирации – состоит из β-инспираторных нейронов и инспираторно-тормозных (ИТ), т.е. экспираторных нейронов. β-инспираторные нейроны возбуждаются афферентными сигналами от РРЛ. На механизм выключения вдоха влияют также нисходящие импульсы от ПТЦ.
Хеморецепторы возбуждаются постоянно и постоянно посылают импульсы в продолговатый мозг, возбуждая α-инспираторные нейроны. Они возбуждаются и посылают импульсы к мотонейронам спинного мозга.
Мотонейроны возбуждаются и посылают импульсы к мышцам. Они сокращаются и наступает вдох.
При вдохе объем легких увеличивается и возбуждаются РРЛ, которые посылают возбуждающие импульсы по чувствительным волокнам блуждающего нерва к β-инспираторным нейронам. В результате суммации импульсов от α-инспираторных нейронов и рецепторов растяжения легких достигается порог и возбуждаются и β-инспираторные нейроны благодаря влиянию вышележащих отделов ЦНС.
Инспираторно-тормозные нейроны посылают тормозные импульсы к α-инспираторным нейронам. В результате α-инспираторные нейроны тормозятся и не посылают импульсы к мотонейронам. Мышцы расслабляются, происходит выдох.
К β-инспираторным нейронам не поступают импульсы, и они тормозятся (не возбуждаются).
β-инспираторные нейроны не возбуждают инспираторно-тормозные нейроны и поэтому они не посылают импульсы к α-инспираторным нейронам. α-инспираторные нейроны вновь возбуждаются импульсами от хеморецепторов и наступает вдох.